Why Choose Us
Our Product
The product range covers a variety of inhibitors, agonists, and compound libraries, more than 10,000 specific inhibitors, agonists, and Libraries of over 100 active compounds.
Product Application
Is committed to providing novel and complete high quality small molecule active compounds to customers worldwide.
Our Advantages
We insist the principle of "Quality first, customer foremost, best credit, be pioneering and creative", looking forward to establishing good business relationships with customers all over the world.
Customer Service
Is to be recognized by our clients as one of the best suppliers of chemical products for our superior quality, efficient and cost-effective services.
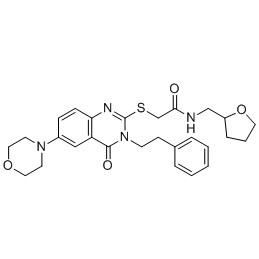
What is Proteasome/Ubiquitin Library?
The Proteasome/Ubiquitin Library refers to a comprehensive collection of reagents and tools that are used to study the ubiquitin-proteasome system (UPS), a major pathway for protein degradation in eukaryotic cells. This system is responsible for the targeted degradation of proteins that are no longer needed, misfolded, or potentially harmful to the cell.



Working Principle Of Thymo Proteasome/Ubiquitin Library
Recently, we identified a novel catalytic subunit designated β5t that is expressed exclusively in cortical thymic epithelial cells (cTECs); these cells are responsible for positive selection of developing thymocytes. The mechanism underlying the thymus-specific expression of β5t is not known. β5t is structurally related to β5 and β5i and is encoded by an intronless gene, in contrast to the three-exon β5 and β5i genes.
Whereas the chymotrypsin-like activity of proteasomes is thought to be important for production of peptides with high affinities for MHC class I clefts, incorporation of β5t into proteasomes instead of β5 or β5i selectively reduces this activity. Although this characteristic differs from the imunoproteasome, the high overall sequence similarity suggests these subunits belong to the same family. β5t and the immunosubunits β1i and β2i, but not their standard counterparts, are incorporated into a vertebrate-specific alternative 20S proteasome referred to as the ‘thymoproteasome’.86) We found that β5t-deficient mice displayed major (but not total) defects in the thymic development of CD8+ T cells, although no obvious abnormality was observed in the thymic architecture, suggesting that β5t is not essential for the differentiation and proliferation of cTECs. Importantly, no obvious alterations in the CD4+CD8+ (co-receptors of TCR) double-positive thymocytes and CD4+ T cell populations were observed in β5t−/− mice. These results suggest a key role for β5t in the development of the MHC class I-restricted CD8+ T cell repertoire during thymic selection.
During positive selection, double-positive cells that interact with self-peptide–MHC complexes expressed on cTECs with a sufficiently modest avidity [i.e., affinity (MHC-TCR interaction) X density (surface MHC levels)] are rescued from intrathymic death and induced to differentiate into CD4+ or CD8+ single-positive thymocytes. In contrast, double-positive cells that interact with high avidity with self-peptide–MHC complexes are eliminated through apoptosis, a process referred to as negative selection.87)–89) In addition, thymocytes that lack functional T cell receptors also undergo apoptosis, a process referred to as null selection. To date, however, the mechanism by which cTECs provide the specialized signals for positive selection has not been elucidated. Considering that proteasomes are essential for the production of MHC class I ligands and that β5t specifically attenuates the proteasomal chymotryptic activity without changing the caspase- and trypsin-like activities, it is possible that thymoproteasomes in cTECs predominantly produce moderate avidity MHC class I ligands rather than high-affinity ligands, which would support positive selection. The discovery of the thymoproteasome may contribute to our understanding of how positive selection occurs in the thymus. The types of antigenic peptides generated by the thymoproteasome, the underlying mechanism, and the roles in positive selection require further examination.
Like the immunoproteasome genes, the β5t and PA28α/β genes appear to result from modification and duplication of existing nonimmune genes, such as β5 and PA28γ, respectively, and may have been instrumental in the emergence of the adaptive immune system.

Substrates to be degraded are marked with ubiquitin, a small (76-amino acid) globular protein, by the concerted action of ubiquitin-activating enzymes (E1), ubiquitin-transferring enzymes (E2) and ubiquitin ligases (E3) (7). Ubiquitin can be ubiquitylated on any of seven lysine (K) residues or the N-terminal methionine (M), leading to eight structurally distinct types of polyubiquitin chains (M1-, K6-, K11, K27-, K29-, K33-, K48- and K63-linked chains) with various lengths. Among them, K48-linked chains are most abundant and play an essential role in proteasomal degradation. Ubiquitylation, like phosphorylation, also functions as a signal in DNA repair, protein trafficking, and NF-κB activation. Monoubiquitylation or K63- and M1-linked polyubiquitylation regulates protein interactions and enzyme activity in proteasome-independent processes, but recent studies have revealed that such chains also play proteolytic roles. Removal of these signals is achieved by deubiquitylating enzymes (DUBs), which antagonize the ubiquitylation machinery by removing ubiquitylation modifications from their substrates. Effector proteins harbouring ubiquitin-binding domains (UBDs) are assumed to function as readers/decoders by discriminating specific chain types. In addition to homotypic chains, cells contain heterotypic ubiquitin chains in which multiple linkages form mixed or branched chains.
The 26S proteasome is a 2.5-MDa complex of 33 different subunits, which are arranged into an elongated structure composed of a central 20S core particle (CP) with one or two terminal 19S regulatory particle(s) (RP). The CP consists of two outer α rings and two inner β rings, which are composed of seven structurally similar α and β subunits, respectively. Three of the seven β subunits have proteolytic sites (β1, β2, and β5; caspase-like, trypsin-like, and chymotrypsin-like activities, respectively) on the inner surface of the chamber formed by two abutting β rings. The combination of multiple active sites with different selectivity contributes to rapid and processive degradation of substrates that have entered the catalytic chamber. In mammalian cells, the proteolytic subunits can be replaced with immunotype subunits to form tissue-specific proteasomes, called the ‘immunoproteasome’ and ‘thymoproteasome’ that generate specific peptides for antigen presentations. The central narrow channel of the α ring forms an entrance gate for substrates, which is essentially closed in CP alone. Association of the RP or other proteasome activators such as PA28α/β (encoded by PSME1/2), PA28γ (PSME3) and PA200 (PSME4) induces gate opening, converting the CP into the degradation-competent state. Because the translocation channel is very narrow, a substrate protein must be unfolded before it can enter the channel. Hence, even in the open state, well-folded proteins do not enter the chamber of the CP. Accordingly, the proteasome is referred to as a ‘self-compartmentalizing protease’ (14). The RP consists of six ATPase subunits (Rpt1–6) and 13 non-ATPase subunits (Rpn1–3, 5–13 and 15), which play roles in processing ubiquitylated substrates. The ubiquitin receptor subunits (Rpn10, Rpn13 and the recently characterized Rpn1) capture ubiquitylated substrates directly or indirectly, whereas the ATPase subunits form a ring structure that promotes substrate unfolding and translocation into the CP, driven by ATP hydrolysis. Because PA28α/β, PA28γ, and PA200 do not utilize ATP and ubiquitin, the substrates of these alternative activators are generally unstructured proteins or peptides.
The Structure Of The Ubiquitin Library–Proteasome System
Protein metabolism, pivotal for normal cellular function, involves both synthesis and degradation of proteins on a constant basis. Eukaryotic cells are equipped with three different systems to accomplish protein degradation: the mitochondrial proteases, which degrade the majority of mitochondrial proteins, the lysosomes, which degrade membrane and endocytosed proteins, and the ubiquitin–proteasome system, which degrades the vast majority of long- and short-lived normal and abnormal intracellular proteins. In fact, up to 80–90% of all intracellular proteins are degraded via the ubiquitin–proteasome system, which is hence considered to be the major pathway of intracellular protein degradation and of utmost significance for cell biology.
Degradation of proteins via the ubiquitin-proteasome system involves two distinct, sequential steps. In the first series of reactions, multiple moieties of ubiquitin are activated, transferred, and bound to cellular proteins by action of E1, E2, and E3 enzymes, respectively, generating a polyubiquitin chain, which sometimes requires the stabilizing action of an E4 molecule. In the second step, cellular proteins, labeled by a chain of at least four ubiquitin moieties, are recognized and degraded by the 26S proteasome complex. Of note, the proper interaction of some proteins with the ubiquitin system or the proteasome complex necessitates the facilitating action of chaperones such as HSP70 or chaperone-like molecules such as the valosin-containing protein.
Illustration of the ubiquitin–proteasome system. The ubiquitin system comprises three sucessive steps: the activation of ubiquitin (Ub) by the formation of a thio-ester binding to E1 (Ub activating enzyme), the tranfer of Ub via E2 (Ub carrier enzyme) to E3, which covalently binds Ub to a lysine residue of the target protein either directly (RING domain E3) or via an intermediate thio-ester formation within the E3 (HECT domain E3). This process, leading to mono- or polyubiquitination, is balanced by the enzymatic process of de-ubiquitination, mediated by a number of enzymes. Whereas mono-ubiquitination exerts a modulatory effect upon protein function, poly-ubiquitination leads to protein degradation. Facilitated by the action of carrier molecules such as valosin-containing protein (VCP) in certain cases, proteins, tagged with a chain of at least four Ub molecules, bind to the recognition site on the 26S proteasome complex. The 26S proteasome complex comprises a central 20S proteolytic core, formed by four rings in barrel-shape structure, and two 19S regulatory units to either side of it. Besides their recognition function, the 19S regulatory units are mainly responsible for unfolding the substrate proteins into the direction of the 20S proteasome. Passage through this proteolytic tunnel, mainly the inner beta-rings, which harbor most of the proteolytic activity, leads to the generation of protein fragments, six to eight amino acids in length. Detachment of VCP and the Ub chain from the target protein prior to its move through the proteolytic tunel restores their intracellular pools.
The Regulation Of The Ubiquitin Library–Proteasome System

For fine-tuning of cellular function, the ubiquitin–proteasome system is regulated tightly and with a high degree of specificity. This is accomplished primarily on the level of the E3 enzymes, which finalize the ubiquitination cascade by the formation of an isopeptide bond between the activated C-terminal glycine residue of ubiquitin and an amino group in position of a lysine residue of either the target protein or a previously conjugated ubiquitin moiety following association with the target protein on the basis of certain recognition patterns. One of the most important recognition patterns is a “destabilizing” (free basic, bulky hydrophic, or uncharged) N-terminal amino acid. This sequence can be a standard characteristic of a protein (primary destabilizing N-terminal amino acids, e.g. arginine and lysine) or may be generated via one (secondary destabilizing N-terminal residues such as apartate, glutamate, and cystein) or two (tertiary stabilizing N-terminal residues such as asparagine and glycine) intermediate steps.
Clearly, this system relates the half-life of an intracellular protein to its N-terminal residue, which has become known as the N-end rule. It also mediates degradation of proteins after their initial cleavage, for instance cohesin, which is involved in the cohesion of sister chromosomes during mitosis. The overall significance of the hierarchical system of the N-end rule pathway for the development of the cardiovascular system has recently been demonstrated in the severe heart defects and the impairment of maturation of early vascular plexus occurring as a consequence of ATE-1-targeted deficiency in Arg-tRNA-protein transferases, which mediate the conjugation of arginine to the N-termini of proteins as part of the pathway. Other important substrate recognition patterns for the ubiquitin–proteasome system are protein phosphorylation and hydroxylation. Proteins that undergo ubiquitin-mediated degradation following phosphorylation or hydroxylation include the inhibitory molecules to nuclear factor kappa B (IκB) and hypoxia-inducible factor 1 alpha (HIF-1α), respectively.
Recently, it was discovered that the consequences of polyubiquitination are crucially determined by the position of the ubiquitin lysine residue involved in the reaction. Whereas polyubiquitination involving the lysine residue at position 48 of the amino acid sequence of ubiquitin leads to protein degradation by the proteasome, modulation of protein function remains to be the only consequence of polyubiquitination involving the lysine residue at position 63. An intriguing example for this latter aspect is the regulation of the IκB kinase complex (IKK) by so-called K63 polyubiquitination, which seems to be additive and even complementary to IKK activation by phosphorylation. In addition to K63 polyubiquitination, monoubiquitination has been described as yet another mode of functional regulation of the activity of proteins, e.g. in the metabolism of cell surface receptors.
In addition to its substrates, the activity of the ubiquitin system itself can be modulated by a number of factors, including glucocorticoids, thyroid hormones, cytokines, and cancer-expressed proteins such as proteolysis-inducing factor (PIF). Notably, some factors, e.g. interferon gamma (IFNγ), not only stimulate the modification of substrates of the ubiquitin system, including the IκB family of proteins, but, furthermore, modify the components of the enzymatic machinery of the ubiquitin system and the proteasome complex. Further coordination between substrates and system might be reflected by the fact that protein kinase C and tyrosine kinase pathways are not only involved in the modifcation of substrates of the ubiquitin–proteasome system but also mediate the phosphorylation of E1 and E2, thereby increasing their activities by even more than twofold.
Certifications

Our Factory
We insist the principle of "Quality first, customer foremost, best credit, be pioneering and creative", looking forward to establishing good business relationships with customers all over the world. Suzhou Ureiko is a reliable world supplier in fine chemicals and custom manufacturing. We understand your needs, and will try to help our customers to advance the research and development in a wide variety of areas. We strive to benefit our customers and partners.Let's work together for bright future!

FAQ
As one of the most professional proteasome/ubiquitin library enterprises in China, we're featured by small molecules chemicals and inhibitors. Please rest assured to buy custom made proteasome/ubiquitin library from our factory. For OEM service, contact us now.